

А.А. Осипова[1]
Факультет фундаментальной медицины Московского государственного университета имени М.В.Ломоносова, отделение «Фармация», Москва, Россия
E-mail: alinaosipov@mail.ru
[1] ORCID: 0000-0003-2846-8230
Аннотация. В статье рассматривается роль и функции лептина в регуляции энергетического обмена и функционировании организмов живой природы.
Ключевые слова: лептин, функции организма, энергетический обмен
1. Введение
Ob модель ожирения впервые была описана на мышах в 1950 г. Тучный фенотип передаётся аутосомно-рецессивным методом. Эксперименты показали, что ob мыши страдают генетическим расстройством, приводящим к неэффективной продукции гуморального фактора, снижающего питание в ответ на ожирение. В 1994 году последовательность ob гена была описана. Белок – продукт гена был назван лептином от греческого слова leptos (тонкий). Именно мутации в этом гене, приводящие к гиполептинемии, могут быть одной из причин ожирения. Лептин играет роль и в других важных процессах, например, регулируя работу репродуктивной системы и участвуя в кроветворении [10]. Открытие лептина доказало тот факт, что жировая ткань является сложным гормонально активным органом, играющим важнейшую роль в регуляции функционирования организма [13]. Было обнаружено, что секретируемые жировой тканью вещества – адипокины – обладают разнообразными эффектами. Оказывая воздействие аутопаракринным способом, они регулируют работу адипоцитов. Поступая в кровь, адипокины влияют на функции различных органов и систем организма: мозг, печень, мышцы, почки, эндотелий, иммунную систему и др. Кроме того, было обнаружено, что жировая ткань экспрессирует ряд рецепторов, позволяющих ей реагировать на афферентные сигналы из эндокринных органов и нервной системы. Многие гормоны – катехоламины, инсулин, кортикостероиды, андрогены и др. – в свою очередь оказывают влияние как на функцию адипоцитов, так и на эффекты адипокинов [5,7,8,14].
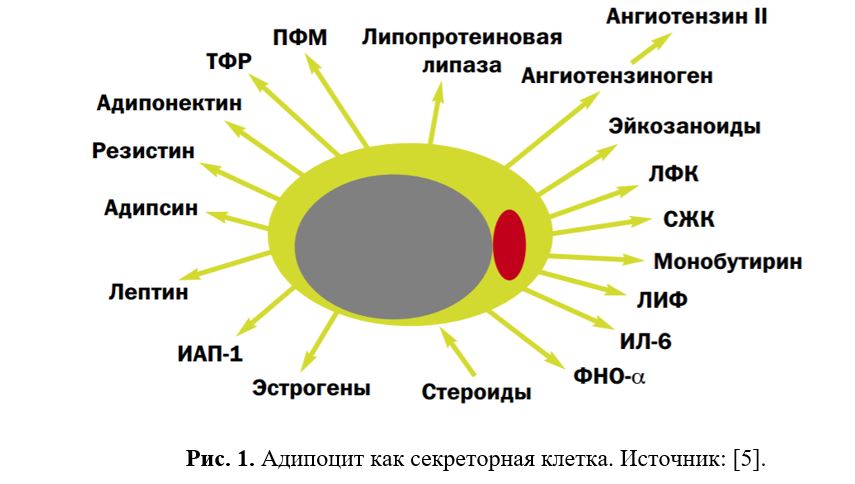
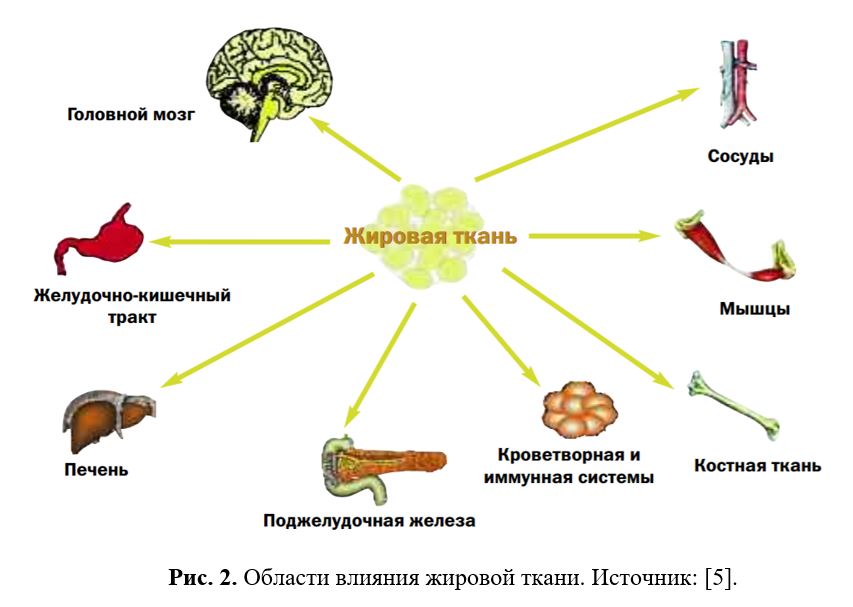
Таким образом (см. Рис.1 и 2), помимо депонирования энергии, жировая ткань через адипокины обладает способностью взаимодействовать с различными органами и системами, включая ЦНС, тем самым регулируя функции организма, а через взаимодействие с нейроэндокринной системой участвовать в адаптации организма к различным внешним воздействиям, таким как, например, голод и стресс [1,5,15].
2. Структура лептина
Лептин – мономерный белок (по разным данным, содержащий 145, 146 или 167 аминокислотных остатков), экспрессирующийся почти исключительно адипоцитами. Молекулярная масса составляет 16 кДа. По пространственной структуре он относится к группе альфа-спиральных белков, в которую входят также гормоны роста, пролактин и цитокины.
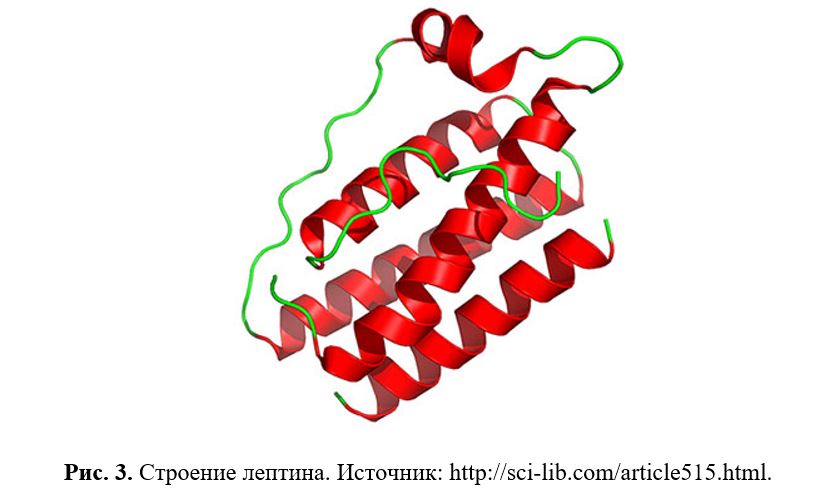
4 антипараллельные α-спирали (A, B, C, D) с восходяще-нисходящим соединением связаны одной короткой (BC) и двумя длинными (AB и CD) петлями. Лептин имеет два консервативных цистеиновых остатка (один в CD-петле и C-концевом остатке), которые образуют дисульфидный мостик, соединяющий петлю CD с C-концевой частью D спирали (Рис. 3). Дисульфидный мостик имеет решающее значение для его структурной стабильности, секреторной и биологической активности. Также данные структурные характеристики напоминают обнаруженные в общем в цитокинах: гранулоцитарно-колониестимулирующем факторе (G-CSF) и интерлейкине-6 (IL-6), так что лептин классифицируется как длинноцепочечный цитокин.
2.1 Биосинтез и секреция
Ген, кодирующий продукцию лептина, называется геном ожирения (ob ген), то есть лептин является продуктом его экспрессии. Подавляющее количество лептина секретируется гормонально адипоцитами в белой жировой ткани (подкожный жир) и в небольшом количестве – в бурой (внутренний жир). Паракринная и аутокринная секреция наблюдается в гонадах, гипоталамусе, гипофизе и плаценте. Кроме того, экспрессия гена лептина отмечается в желудке, скелетных мышцах и эпителии молочной железы.
Размер адипоцита является важной детерминантой синтеза лептина: крупные жировые клетки содержат гораздо большее количество гормона, чем мелкие адипоциты.
Адипоциты выделяют лептин в кровь прямо пропорционально массе жировой ткани. Экспрессия и секреция лептина регулируется также посредством многообразия других факторов: увеличивается под влиянием инсулина, глюкокортикоидов, TNF-α, эстрогенов, а снижается посредством β3-адренэргической активности, андрогенов, свободных жирных кислот, гормона роста, грелина [11].
2.2 Транспортные белки
Лептин циркулирует в кровотоке как в свободной, так и связанной со специфическими белками форме. Считается, что короткие формы рецепторов, экспрессирующиеся в хориоидном сплетении, вовлечены в его транспорт и могут облегчать процесс проникновения лептина через гематоэнцефалический барьер для взаимодействия с длинной формой в гипоталамусе, а также участвуют в почечном клиренсе. Также речь идет о растворимом рецепторе лептина.
2.3 Рецепция и проведение сигнала
Как любой гормон, лептин проявляет свои эффекты на организм через рецепторы. Лептиновый рецептор кодируется геном диабета (db ген). Рецепторы лептина – члены суперсемейства рецепторов цитокинов 1-го класса и характеризуются экспрессией как в центральной нервной системе, так и на периферии.
Идентифицировано три различных варианта рецептора:
- Растворимый рецептор лептина;
- Связанный с мембраной рецептор лептина, который имеет короткий внутриклеточный домен и не способен осуществлять трансдукцию гормонального сигнала;
- Связанный с мембраной рецептор, имеющий длинный внутриклеточный домен и способный передавать гормональный сигнал.
В то же время, известно не менее 6 изоформ рецепторов лептина: obRa, obRb, obRc, obRe, obRf, obRa, obRc, obRf принадлежат к коротким формам, а obRe является растворимой.
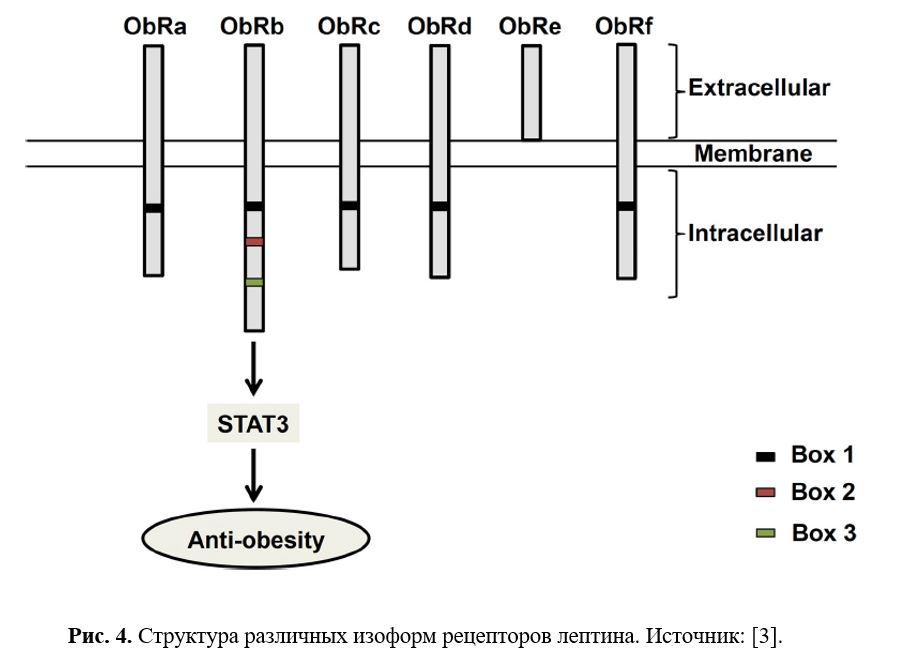
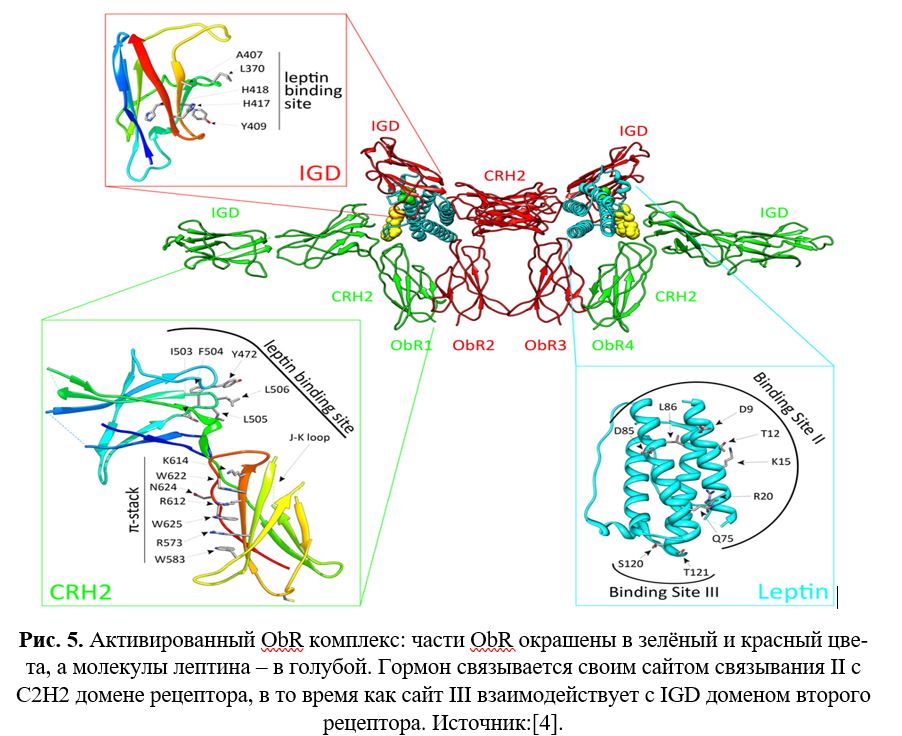
Все изоформы имеют идентичную внеклеточную часть, состоящую из шести доменов: N-концевого домена (NTD), двух доменов CRH (CRH1 и CRH2), иммуноглобулиноподобного домена (IGD) и двух дополнительных мембранно-проксимальных фибронектинов типа III (FN III) (Рис. 4).
Из числа различных видов коротких изоформ лучше всего изучена obRa. Наибольшая концентрация рецептора отмечена в почках (в мезангиуме и сосудах).
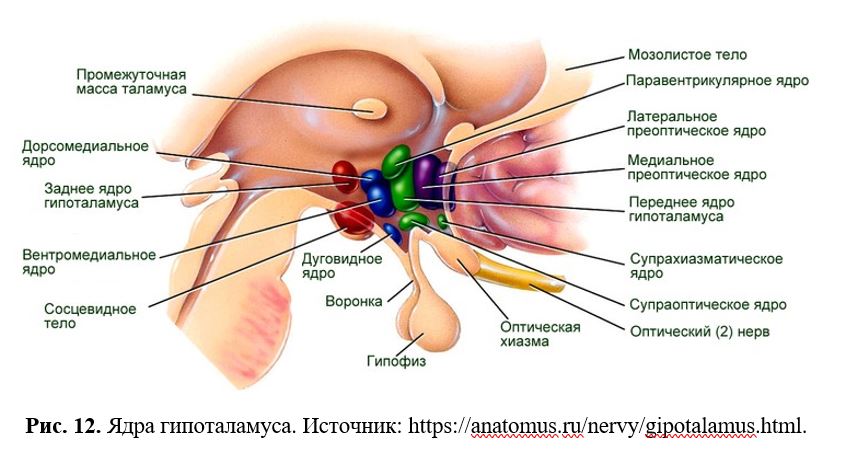
Полнофункциональной является единственная удлиненная форма obRb, через которую и осуществляется действие лептина. У человека и животных данные рецепторы были обнаружены в гипоталамусе, надпочечниках, поджелудочной железе и жировой ткани. Эта форма рецептора лептина с длинным цитоплазматическим доменом наиболее активно экспрессируется в гипоталамусе и имеется, главным образом, в аркуатном ядре (ARC) и вентромедиальном ядре (VMH), а также в паравентрикулярном ядре (PVN), латеральном гипоталамусе (LH), вентральном премамиллярном ядре (PMv) и дорсомедиальном ядре (DMN).
Механизм проведения сигнала рецепторов цитокинов включает в себя активацию ассоциированных с рецептором тирозинкиназ, фосфорилирование рецептора этими киназами с формированием сайтов посадки для ряда белковых эффекторов и фосфорилирование некоторых из них теми же тирозинкиназами. Рецепторы лептина взаимодействуют с внутриклеточными мессенджерами: киназой митоген-активированного белка (MAP), инсулин-рецепторным субстратом (IRS-1; IRS-2), Janus-киназой, оксидом азота (NO) и сигнальным трансдуктором и фактором активации транскрипции (STAT).
ObRb содержит последовательности, которые определяют взаимодействие цитоплазматического домена с киназой Януса и белками активаторами транскрипции.
ObRb активация зависит от CRH2 (cytokine receptor homology domain), IGD (иммуноглобулиноподобного) и FN (фибронектинового) III доменов. Домен CRH2 является определяющим фактором связывания лептина. Область из четырех последовательных гидрофобных остатков была идентифицирована как сайт связывания лептина в CRH2 домене. IGD и проксимально-мембранные домены не имеют сродства к лиганду, но, тем не менее, необходимы для активации рецептора. Лептин содержит сайт связывания III, который делает возможным контакт с IGD со второй частью ObRb, чтобы произошла димеризация и последующая активизация. В FN III доменах содержится два консервативных цистеина, которые требуются для активации рецепторного комплекса (Рис. 5 и 6).
До взаимодействия с лептином рецепторы пребывают в форме мономера. После прикрепления гормона формируется димер-активный гормон-рецепторный комплекс, который проникает в ядро и оказывает эффекты на экспрессию соответствующих генов.
При малой концентрации гормона (число молекул гормона много меньше числа рецепторов) он связывается с двумя молекулами рецептора, позволяя их внутриклеточным доменам связать Jak-киназы, которые начинают фосфорилировать друг друга, активируя передачу сигнала. Однако при равной концентрации агониста и рецептора передача сигнала по такому механизму невозможна, так как каждый рецептор связан с лигандом, что препятствует их димеризации.
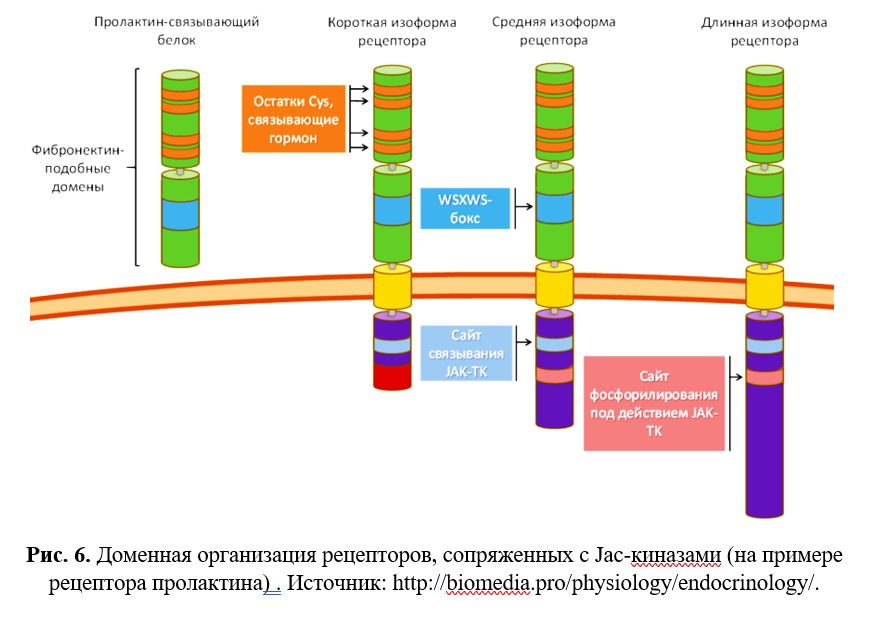
Структурно каждый мономерный рецептор, ассоциированный с тирозинкиназой класса Janus, состоит из 3 частей:
- Внеклеточная часть, связывающая гормон, содержащая:
- Остатки Cys, связывающие лиганд
- WSXWS-бокс, обеспечивающий свёртывание, транспортировку и димеризацию
- Трансмембранная гидрофобная часть
- Внутриклеточная часть, содержащая:
- Участок связывания Jak-киназ
- Участок фосфорилирования Jak-киназами (за наличие во внутриклеточной части данных противоположных доменов эти соединения назвали Janus-киназами — в честь древнеримского бога двуликого Януса).
В целом, рецепторы, ассоциированные с тирозинкиназами класса Janus, передают сигнал по нескольким сигнальным каскадам: STAT, МАР-киназному и фосфатидилинозитольному путям.
2.4 STAT-путь (англ. «Signal Transducers and Activators of Transcription»)
STAT белки – это семейство факторов транскрипции эукариот, которые участвуют в передаче сигнала от большого числа цитокинов и факторов роста.
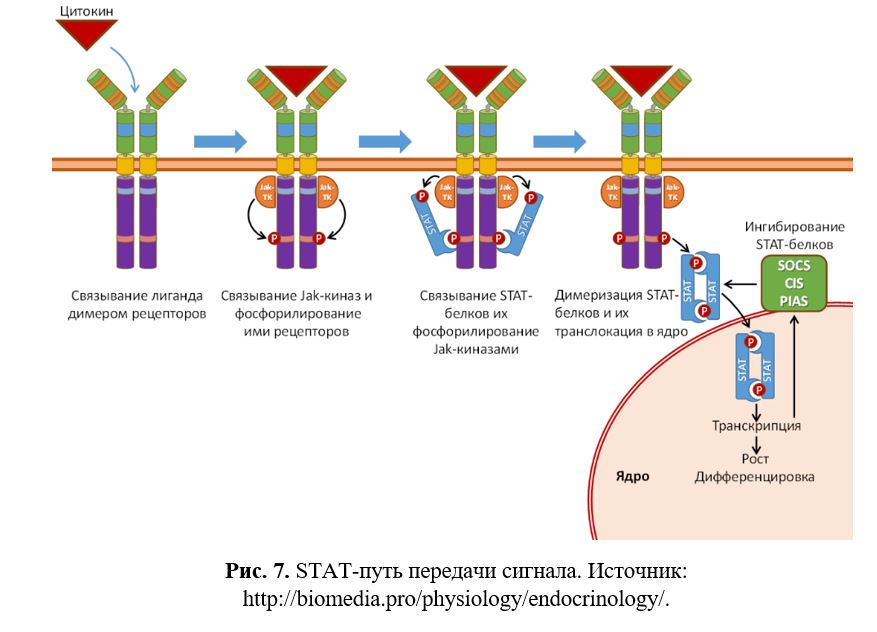
Активированные Jak-киназы передают сигнал по STAT-пути {рис.8}. После связывания гормона, димеризации рецепторов и фосфорилирования их внутриклеточных доменов Jak-киназами на них садятся STAT-белки. STAT-белки содержат SH2 домен, который связывается с фосфотирозином рецептора. Под действием Jak-киназ они претерпевают активирующее фосфорилирование, после чего диссоциируют от рецептора направляются в ядро, где активируют транскрипцию генов-мишеней и генов, кодирующих ингибиторы STAT-пути (белки SOCS, CIS и PIAS).
Активация происходит за счёт того, что димеры STAT взаимодействуют в этих генах со специфическими, как правило, полинуклеотидными последовательностями TTCCGGGAA. Каждый цитокин активирует характерный для него набор STAT и, следовательно, индуцирует характерный именно для него набор генов (Рис. 7). Сложность данного процесса возрастает в виду того, что участники разных путей взаимодействуют друг с другом, образуя целые сети передачи сигналов.
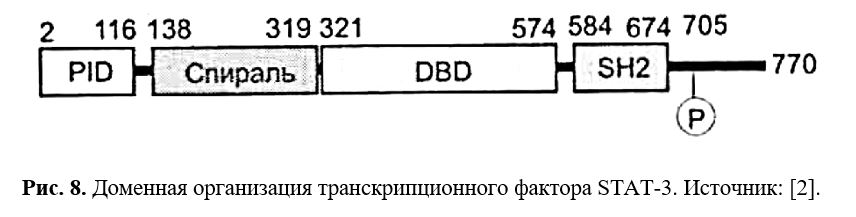
Если говорить точнее, то в случае лептина при активации JAK2 консервативный ObRb фосфотирозиновый 1138 мотив служит сайтом связывания для SH2 домена STAT3. Одновременно лептином активируется STAT-1, STAT-5 и STAT-6 пути, однако in vivo наблюдали лишь STAT-5 фосфорилирование (Рис. 8).
Кроме того, ObRb лептина имеет три консервативных остатка тирозина, где также имеется сайт связывания SH2 домена. Фосфорилирование SH2 через JAK в конечном итоге приведет и к активации MAPK (активированной митогеном протеинкиназой) ERK 1,2 (внеклеточной сигнальной киназы 1,2). Фармакологическое ингибирование ERK 1,2 приводит к блокированию анорексигенных эффектов лептина в гипоталамусе. Кроме того, также была продемонстрирована важная роль PI3K (фосфатидилинозитольно-киназного пути) в передаче сигнала от ObR при введении инъекций с ингибиторами PI3K. В данном случае анорексигенный эффект лептина также блокируется. Но этот путь является общим с другими рецепторами, поэтому сложно оценить ObR-специфичный вклад PI3K в поддержание энергетического гомеостаза.
3 Эффекты, функции и связи лептина
Основной функцией лептина является обеспечение баланса между жировыми запасами и расходованием и потреблением пищи. Лептин оказывает два ряда эффектов: тормозит пищевое поведение и стимулирует сжигание жиров в энергообмене. Анорексический эффект воспроизводится N-концевым фрагментом лептина 1-35: он стимулирует центр насыщения, угнетая секрецию центра аппетита и орексигенного NPY (нейропептида Y). В рамках целого организма это приводит к торможению пищевого поведения и активации центра теплопродукции.
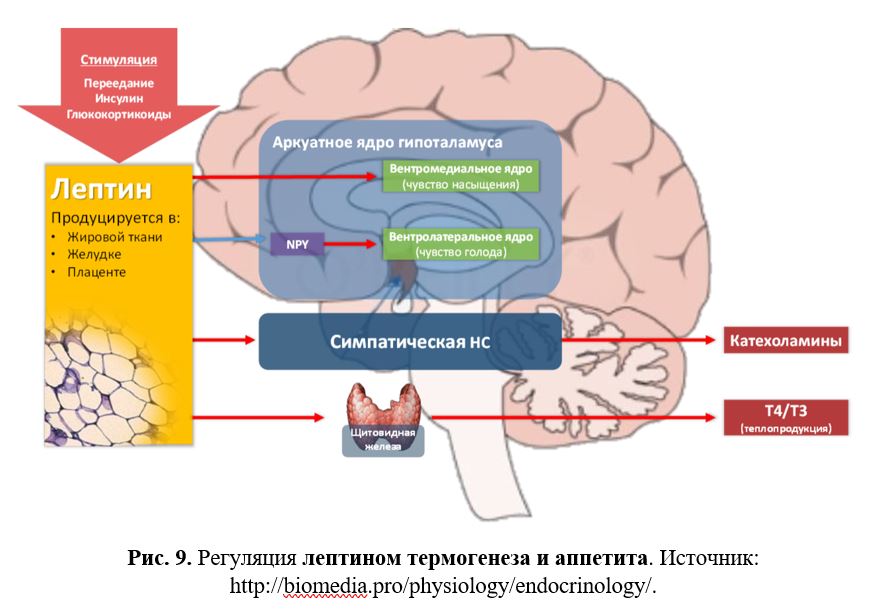
Существенно, что эффективность лептина сохраняется при его введении в желудочек мозга, т.е. основной мишенью лептина является центральная нервная система, преимущественно гипоталамические структуры. Стимул секреции лептина адипоцитами — приём пищи (Рис. 9). Этот быстрый эффект опосредован, по-видимому, повышением секреции инсулина и глюкокортикоидов. Негативный регулятор секреции лептина – голодание и стимуляция рецепторов-активаторов пролиферации пероксисом PPAR. Выявлен ещё один механизм действия противодиабетических препаратов – торможение через PPARγ образования адипоцитами TNFα и лептина, оказывающих липолитическое действие (в частности, антидиабетическим препаратом троглитазоном).
Кроме того, лептин является главным стимулятором экспрессии CART в гипоталамусе. CART (cocaine-amphetamine regulated transcript) – семейство пептидов транскрипта, регулируемого кокаином и амфетамином, — нейромедиатор/нейромодулятор и, возможно, паракринный фактор, поскольку перифирическое введение даже в высоких дозах не оказывает действия. Центральное введение CART оказывает мощное анорексигенное действие и интенсификацию энергообмена. Описан ряд мутаций генов лептина (линия мышей ob/ob) и его рецепторов (линия мышей db/db, линия крыс Zucker, мутация fa); мутации сопровождаются сходными симптомами – ожирением, гиперфагией, сниженным энергообменом (гипотермия, пониженное потребление кислорода) и физической активностью, резистентностью тканей к инсулину при гиперинсулинемии и гипергликемии.
Мутации лептина и его рецептора – причина небольшой части случаев ожирения человека.
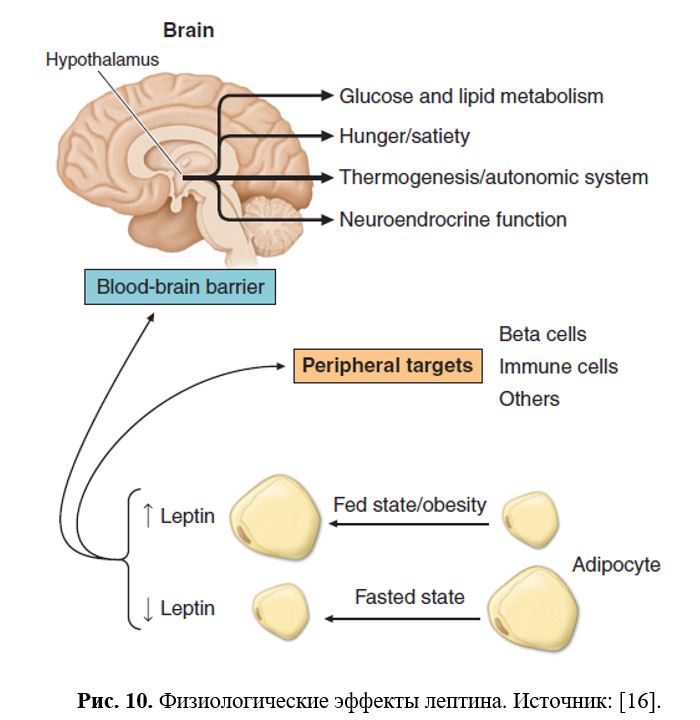
Помимо регуляции энергетического баланса, лептин опосредует нейроэндокринную адаптацию к голоданию, в частности, изменения репродуктивной и тиреоидной функций. Лептин служит пермиссивным фактором в отношении полового созревания (Рис. 10).
Обобщая функции лептина, можно отметить следующие:
- Анорексигенное действие
- Угнетение пищевого поведения и снижение активности центра аппетита
- Стимуляция энергетического обмена
- Активация центра теплопродукции
- Утилизации жиров и снижение массы тела
- Индукция начала и увеличение скорости полового созревания
- Поддержание репродуктивной функции
- Стимуляция секреции Гн-РГ, ЛГ и ФСГ
- Регуляция функций тиреоидной оси
- Адаптация функций щитовидной железы к голоданию
- Снижение синтеза и секреции NPY в мозге
- Стимуляция синтеза и секреции КРГ
- Стимуляция секреции грелина
- Стимуляция секреции адипонектина
- Снижение секреции инсулина
Перейдем к более детальному рассмотрению.
4 Лептин и ожирение
4.1 Ожирение
Ожирение — избыточное отложение триглицеридов в адипоцитах — увеличивает риск смерти человека репродуктивного возраста (20–50 лет) на 50%. Ожирение способствует развитию сердечно-сусудистых заболеваний, сахарного диабета, желчно-каменной болезни, цирроза печени, рака эндометрия и молочной железы (из-за увеличенной ароматизации андрогенов в эстрогены в адипоцитах).
Различают детское и взрослое ожирение. В первом случае к ожирению ведет увеличение количества адипоцитов, вызываемое усиленной дифференцировкой преадипоцитов в адипоциты под действием избыточного потребления/образования жирных кислот в детском возрасте.
Одним из сигналов дифференцировки служит активация жирными кислотами PPARγ. Взрослое ожирение связано только с гипертрофией адипоцитов. С точки зрения энергетики, подтверждаемой данными практической медицины, ожирение возникает из-за нарушения баланса между потреблением пищевых продуктов и их расходованием на образование тепла и выполнение физиологической и физической работы.
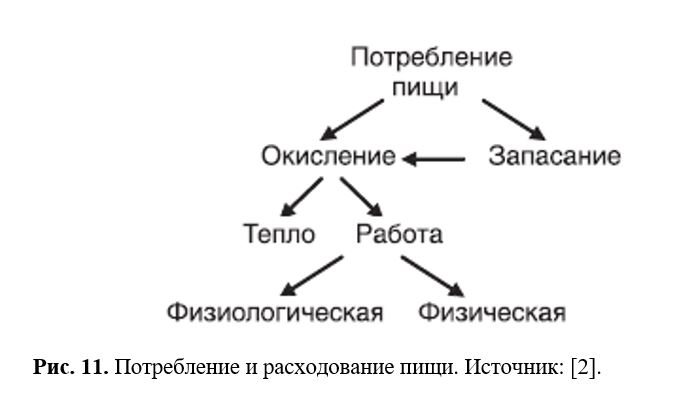
Дисбаланс может возникнуть вследствие нарушений на любой из стадий. Наиболее частой причиной ожирения служит несоответствие потребления пищи (ее количества и качества) и физической активности (Рис. 11). Однажды возникнув, избыток жиров способствует поддержанию нового состояния, в частности, за счет модуляции уровня секретируемого лептина: попытка похудания сопровождается снижением секреции лептина, оказывающего анорексигенное и энергозатратное действие. Аналогичным образом снижение уровня жирных кислот ведет к увеличению секреции инсулина, способствующего увеличению жировых запасов. Как уже упоминалось, к ожирению может вести разрушение у лабораторных животных вентромедиального гипоталамуса, служащего центром «насыщения». В результате развиваются гиперфагия и снижение энергозатрат. К аналогичным последствиям с преобладанием компонентов потребления пищи или расходования энергии ведут инактивирующие мутации системы возникновения/проведения
анорексигенных сигналов (например, мутации генов лептина и его рецептора, ob/ob и db/db соответственно) или активирующие мутации системы орексигенной стимуляции (например, «желтое» ожирение, или ожирение агути).
Дисбаланс между потреблением пищи и энергозатратами может возникнуть и на периферии, за счет звеньев запасания, окисления и сопряжения окисления с синтезом и расходом макроэргов. Одним из примеров может служить ожирение при гипотиреозе или инактивирующих мутациях Т-рецепторов. К настоящему времени выявлены мутации/полиморфизмы многих десятков генов, ассоциированные с ожирением. В силу полифакторности ожирения его рассматривают не как болезнь, а как симптом.
4.2 Гипоталамическая регуляция аппетита
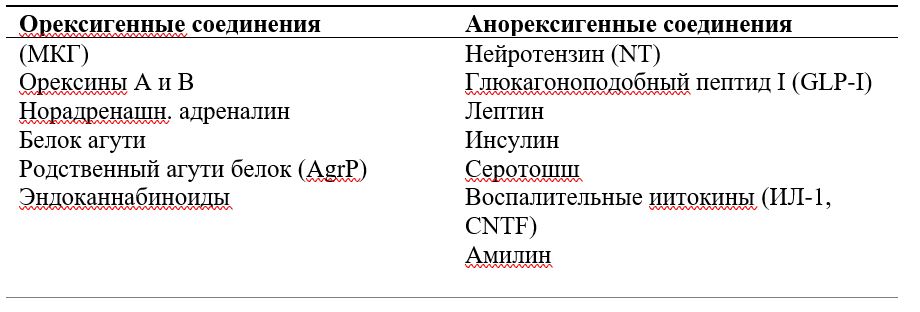
В регуляции пищевого поведения принимает участие целый ряд структур мозга, но базовые процессы протекают в гипоталамусе, куда поступает информация о внешних раздражителях, с рецепторов желудка, об уровне глюкозы и аминокислот в крови, об уровне гормонов и других сигнальных соединений, связанных с пищеварением и энергообменом. Принятие решения об изменении пищевого поведения и реализация этого решения осуществляются с участием множества сигнальных соединений, которые делятся на две основные группы: стимулирующие и подавляющие пищевое поведение (орексигенные и анорексигенные соединения) (Табл. 1).

4.3 Области мозга, связанные с регуляцией потребления пищи
Аркуатное ядро. (ARC): экспрессирует мРНК многих орексигенных и анорексигенных пептидов. Терминали соответствующих нейронов идут в другие отделы мозга, в частности, те, микроинъекции данных пептидов в которые вызывают изменения в потреблении пищи. Многие нейроны ARC экспрессируют одновременно по меньшей мере два регулирующих потребление пищи пептида (например, NPY и AgrP). Внутри ARC терминали NPY-ергических нейронов обнаруживаются на телах GAL- и β-END-ергических нейронов, терминали GAL-ергических нейронов — на телах β-END-ергических нейронов. ARC не защищено гематоэнцефалическим барьером, что обеспечивает возможность его регуляции соединениями, поступающими через кровь и цереброспинальную жидкость.
Вентромедиальное ядро (VMN) и латеральный гипоталамус (LH): VMN чувствительно к орексигенным и анорексигенным соединениям. Его разрушение ведет к гиперфагии и ожирению («центр насыщения»). Терминали из VMN обнаружены в паравентрикулярном и дорзомедиальном ядрах. Через LH проходит переднемозговой пучок. Разрушение LH вызывает временную афагию, а электоростимуляция или микроинъекции орексигенных соединений — повышение потребления пищи («центр питания»).
Дорсомедиальное ядро (DMN): разрушение DMN слабо увеличивает потребление пищи. Микроинъекции орексигенных и анорексигенных соединений соответственно влияют на потребление пищи.Терминали из DMN обнаружены в VMN и паравентрикулярном ядре. Экспрессия NPY в DMN резко возрастает при разрушении VMN и других типах гиперфагии.
Паравентрикулярное ядро (PVN) и околосводный гипоталамус (PFH): чувствительны к микроинъекциям орексигенных и анорексигенных соединений. Разрушение PVN ведет к гиперфагии. PVN — единственная область гипоталамуса, в которой повышается выделение NPY при голодании.
Супрахиазматическое ядро (SCN): повреждения SCN нарушают регулярность приема пищи. Сигналы о светопериодичности направляются через SCN в ARC, VMN, DMN, LH.
Нейроанатомический субстрат контроля аппетита. Рецептивная область для анорексигенных сигналов (лептина, инсулина, КРГ, урокортина, CART) перекрывает рецептивную область для орексигенных сигналов (Рис. 13).
Недостаточность лептина у мышей ob/ob (мутация гена лептина), нечувствительность к лептину у мышей db/db и у крыс Zucker (fa/fa) (мутации генов рецептора лептина) сопровождаются повышением экспрессии и выделения NPY в гипоталамусе и гиперфагией.
При разрушении VMN развивается гиперфагия. При этом мРНК NPY в ARC и уровень, и выделение NPY в PVN снижены. Но при этом гиперфагия остается зависимой от NPY, поскольку иммунонейтрализация NPY блокирует потребление пищи.
При разрушении VMN микроинъекцией колхицина наступает преходящая (4–5 дней) гиперфагия. Сразу же снижается продукция и выделение NPY в ARC и PVN соответственно. Гиперфагия развивается по меньшей мере отчасти за счет повышения чувствительности к NPY и увеличения рецепторов NPY типа Y1 в гипоталамусе.
Параллельно возникает резистентность к лептину. Кроме того, повышается продукция GAL и возрастает чувствительность к нему.
При химической блокировке норадреналинового пучка, идущего из BS, развивается гиперфагия.
При этом исчезает подъем продукции лептина после еды в ночное время и возрастает ночная секреция NPY.
Гипокретины/орексины. Hcrt1 и Hcrt2, или орексины A и B, продуцируются нейронами в дорзальном и латеральном гипоталамусе и PFH.
Терминали обнаружены в ARC, паравентрикулярном ядре таламуса, преоптической области, ядрах перегородки, BS. Внутрижелудочковое введение этих пептидов, а также микроинъекции в LH, PVN и PFH, но не в VMN и POA, стимулируют потребление пищи. Голодание повышает их экспрессию.
Пищевое действие орексинов, по-видимому, определяется рецептором орексинов типа 2. В ARC терминали с орексинами обнаружены на нейронах, коэкспрессирующих NPY и лептин. Вероятно, орексины стимулируют выделение NPY в PVN. И наоборот, терминали с NPY обнаруживаются на нейронах, экспрессирующих орексины, в LH.
Глюкагоноподобный пептид 1 (GLP-1. ). GLP-1 (7-36)амид возникает при процессинге проглюкагона в кишечнике. Иммунореактивный GLP-1 выявлен в нейронах ARC и PVN. Основная часть GLP-1-содержащих терминалей в гипоталамусе идет из ядра одиночного тракта. Внутрижелудочковое введение GLP-1 голодным животным снижает прием пищи. У сытых животных антагонист GLP-1 стимулирует прием пищи и ожирение. GLP-1 ослабляет действие NPY, что позволяет предполагать участие NPY в опосредовании действия GLP-1. Эффект, видимо, связан с ингибированием постсинаптического действия NPY в PVN. Показана коэкспрессия мРНК GLP-1 и рецептора лептина в нейронах BS. Антагонист GLP-1 блокировал тормозное действие
лептина на потребление пищи. Возможно, GLP-1 служит одним из посредников анорексического действия лептина. Интересно, что при нокауте GLP-1-R изменений в потреблении пищи не наблюдается.
Меланокортин (α-МСГ) и белок агути. . У мышей с синдромом ожирения при летальной мутации агути (AY/a) при созревании развиваются ожирение, гиперинсулинемия, гипергликемия, что связано с действием белка агути через меланокортиновые рецепторы MC4.
α-МСГ, как полагают, ингибирует потребление пищи через те же рецепторы. MC4-R обнаружены во многих отделах мозга, включая PVN, VMN, DMN. Мутации MC4-R ведут к последствиям, сходным с синдромом ожирения агути. Введение агониста MC4-R снимает гиперфагию, вызванную введением NPY или мутацией в локусе агути (AY). Введение антагониста MC4-R ведет к повышению потребления пищи сытыми животными. У мышей ob/ob и у нормальных голодных мышей экспрессия ПОМК в ARC снижена. Введение таким животным лептина восстанавливает экспрессию ПОМК. Нейроны, экспрессирующие ПОМК, содержат рецепторы лептина. По-видимому, потребление пищи тонически подавляется системой лептин-ПОМК-αМСГ-MC4-R.
В ARC обнаружены нейроны, коэкспрессирующие NPY и ART (ген, кодирующий родственный агути белок, AgrP ). Экспрессия AgrP возрастает у мышей ob/ob и db/db. AgrP является сильным антагонистом MC3-R и MC4-R. Возможно, AgrP блокирует тормозное действие α-МСГ в PVN на потребление пищи.
Ещё одним медиатором действия лептина является меланинконцентрирующий гормон (МКГ). Действие МКГ противоположно влиянию α-МСГ. МКГ вызывает усиление аппетита и потребления пищи. Транскрипт, регулируемый кокаином и амфетамином (CART). Внутрижелудочковое введение CART тормозит потребление пищи (Рис. 14). Иммунонейтрализация CART повышает ночной прием пищи. мРНК CART обнаружена в ARC, PVN, SON, VMH, переднем PVN и др. областях. Экспрессия CART при голодании и у fa/fa крыс, и ob/ob мышей снижается в ARC и DMH. Введение лептина мышам ob/ob повышает экспрессию CART в ARC. В PVN обнаружены
NPY-терминали на экспрессирующих CART нейронах. Таким образом, посредником биологического действия лептина является CART, содержание которого увеличивается после введения животным кокаина и амфетамина. Лептин стимулирует экспрессию гена CART и вызывает увеличение содержания мРНК CART в гипоталамусе. Эксперименты по введению CART в желудочек мозга крыс выявили уменьшение потребления пищи, стимулированное NPY.
Таким образом, лептин может стимулировать аппетит через подавление экспрессии генов и биосинтеза NPY и МКГ, так и снижать потребление пищи через активацию экспрессии генов ПОМК (МСГ) и CART в нейронах.
Введение лептина подавляет потребление пищи с лаг-периодом 8 – 10 ч. Активная, длинная форма рецептора лептина (OBRb) экспрессируется в ARC, VMH, PVN, LH, вентральном премамиллярном ядре (PMv), DMN. Микроинъекции лептина в ARC, VMH, LH тормозят потребление пищи. В ARC обнаружена коэкспрессия OB-Rb и NPY. Лептин снижает экспрессию NPY в ARC, DMN, PVN. OB-Rb коэкспрессируется в ряде областей с ПОМК, MCH, GAL, орексинами,
GLP-1, что предполагает, что такие нейроны могут быть мишенями для лептина.
У человека циркадный ритм уровня лептина в крови характеризуется базальным уровнем с 8.00 до 12.00 и максимумом между 24.00 и 4.00. Пульсаторный характер с интервалами 44 мин. Циркадные изменения связаны с высотой пульсов. Лептин ингибирует экспрессию гена NPY и секрецию NPY, а также подавляет стимулирующее аппетит действие экзогенного NPY.
Лептин стимулирует экспрессию ПОМК в гипоталамусе и соответственно продукцию α-МСГ. У мышей ob/ob экспрессия ПОМК снижена. Инсулин ингибирует экспрессию NPY и стимулирует экспрессию и секрецию КРГ, который, в свою очередь, индуцирует экспрессию ПОМК и секрецию α-МСГ.
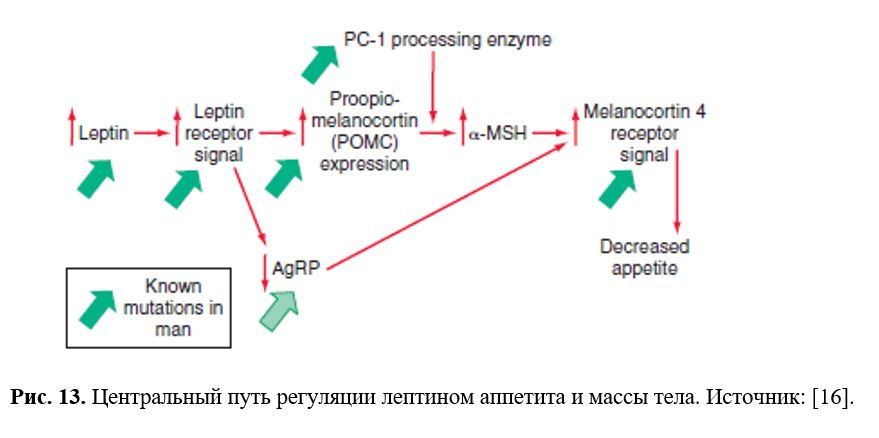
Лептин действует через ПОМК в гипоталамусе, увеличивая продукцию α-МСГ, который, в свою очередь приводит к ингибированию аппетита, влияя на меланокортиновые рецепторы 4 типа. Кроме того, снижается продукция нейропептида AgRP, который в норме выступает антагонистом меланокортиновых рецепторов 4 типа.
Как уже говорилось, адипоциты выделяют лептин в кровь прямо пропорционально массе жировой ткани и состоянию питания. Однако экспрессия и секреция лептина регулируется также посредством многообразия других факторов, что нагляднее всего отражается в таблице (Табл. 2):

Различные биологические эффекты лептина проявляются через его связывание с рецепторами на нейронах. Гормон вызывает снижение аппетита, связываясь с рецепторами на нейронах гипоталамуса, в которых нейромедиаторами являются α-МСГ и NPY. α-МСГ синтезируется в виде предшественника проопиомеланокортина (ПОМК), из которого высвобождается после протеолитического расщепления. Помимо α-МСГ, из ПОМК образуется АКТГ и β-эндорфин.
Лептин тормозит экспрессию гена NPY в нейронах гипоталамуса. Повышенное потребление пищи, вызванное NPY, подавляется лептином. Таким образом, лептин не только снижает синтез NPY, но и тормозит его физиологический эффект. Снижение продукции лептина примерно через сутки после ограничения поступления пищи может быть опосредовано снижением концентрации в плазме инсулина и увеличением концентрации адреналина. Инсулин увеличивает экспрессию гена лептина, тогда как катехоламины снижают её. Спад сочетается с адаптивными физиологическими реакциями на голодание в виде увеличения аппетита и снижения расходования энергии. Такие реакции обычно наблюдались у мышей с недостаточностью лептина и людей с выраженным ожирением. Следует отметить, что терапия малыми дозами приводила к уменьшению гиперфагии и потере массы тела у экспериментальных животных. Напротив, обычные (не связанные с генетическими дефектами) формы ожирения характеризовались повышенным содержанием циркулирующего лептина. Ни эндогенно высокие уровни лептина, ни лечебные мероприятия при помощи введения экзогенного лептина не оказывали влияние на снижение количества жира в организме. Высказано предположение о том, что это связано с развитием лептинорезистентности. Хотя лептин рассматривается как гормон, противодействующий ожирению, при этом состоянии нередко наблюдается гиперлептинемия, как возможное следствие развития лептинорезистентности. Лептинорезистентность может быть следствием дефектов в механизме сигнализации или транспорта лептина через гематоэнцефалический барьер. При лептинорезистентности, наряду с высокой концентрацией лептина в плазме крови, в спинно-мозговой жидкости уровень гормона остаётся минимальным, что позволило высказаться в пользу недостаточного поступления гормона через ГЭБ. Также стало известно, что имеющееся увеличение уровня гипоталамического SOCS3 (супрессор передачи сигналов цитокина) может рассматриваться как молекулярный медиатор приобретенной резистентности к лептину. SOCS3 сдерживает передачу сигналов на рецептор лептина и на другие рецепторы семейства цитокинов, подавляя активность SAK2 и, следовательно, STAT-активации.
Интересно отметить, что гиперфагия и повышение уровня лептина вместе с повышением инсулина являются обычными признаками ожирения. Но в то же время лептин сам по себе является мощным ингибитором потребления пищи и предполагает снижение уровня инсулина путём подавления его секреции и более качественного расходования. Складывается впечатление, что лептин на самом деле не выполняет своей функции как метаболический гормон, ограничивающий избыточную прибавку в весе. Однако видно, что на самом деле лептин не может достигнуть этой цели при гиперлептинемии и лептинорезистентности. Оригинальное объяснение было предложено с позиции «селективной лептинорезистентности», которое основано на сохранении симпатовозбуждающего действия лептина, несмотря на резистентность к потреблению пищи (пресыщению) и на способность гормона снижать массу тела. Если селективная лептинорезистентность имеет место у тучных людей, то лептин может способствовать избыточной симпатической активности и гипертензии, несмотря на резистентность к его метаболическому воздействию.
Сначала было установлено влияние лептина на энергетический метаболизм, связанное с его действием в гипоталамусе. Со временем стало понятно, что действие лептина намного разнообразнее. Кроме ЦНС, он действует на поджелудочную железу, почки, иммунную и симпатическую нервную системы, оказывает влияние на ангиогенез, гематопоэз.
5 Лептин и симпатическая нервная система
Полагают, что симпатическая нервная система является ключевым ингибитором высвобождения лептина. Катехоламины, такие как норадреналин и адреналин, непосредственно ингибируют синтез лептина. В свою очередь, лептин может активизировать симпатическую нервную систему как через гипоталамус, так и путём стимуляции высвобождения адреналина в мозговом слое надпочечников. Такая «раздвоенность» в действии лептина очень важна, так как происходит формирование отрицательной петли обратной связи между лептином и симпатической нервной системой.
6 Лептин и сердечно-сосудистая система
Анализ литературных данных позволяет утверждать, что лептин обладает мощным сосудистым воздействием и участвует в регуляции симпатического тонуса и артериального давления. Регуляторное воздействие на сосудистый тонус и кровяное давление наглядно показано посредством вливания внутривенно или внутрь желудочков мозга животных лептина, что указывает как на центральное, так и на периферическое воздействие гормона. Введение лептина внутрь желудочков мозга вызывает у кроликов повышение артериального давления. Прессорный эффект пропорционален уровню гормона в спинно-мозговой жидкости. Интересно, что доза лептина, необходимая для усиления выброса катехоламинов, по меньшей мере в 100 раз выше для внутривенного, чем для введения внутрь желудочков мозга, что подтверждает взгляд о преобладании центрального механизма в стимуляции симпатической нервной системы и сосудистого тонуса над периферическим. Периферическое воздействие лептина на сосудистый тонус реализуется через некоторые вазоактивные медиаторы, а именно NO и эндотелин-1. Показано, что лептин стимулирует синтез NO и продукцию ЭТ-1 и способствует накоплению активных форм кислорода (АФК) в эндотелиальных клетках пупочной вены у человека. Также известно, что лептин непосредственно индуцирует расширение кровеносных сосудов через NO-независимые пути метаболизма у здоровых мужчин. Таким образом, интегрирующий механизм действия лептина на регуляцию сосудистого тонуса и артериального давления носит, скорее всего, комплексный характер, если учитывать прессорные воздействия (симпатическая активация) и депрессорные воздействия (увеличение NO).
Внутривенное введение лептина животным в течение 90 минут не вызывала увеличения числа сердечных сокращений, в то время как недельная внутрисосудистая инфузия или введение гормона в желудочки мозга вызывали тахикардию параллельно с активацией симпатической нервной системы.
Кроме того, была обнаружена высокодостоверная позитивная корреляция между гиперлептинемией и тахикардией у лиц с избыточной массой тела (за счёт жира) и умеренной гипертензией. Длительная гиперлептинемия может способствовать развитию сердечной недостаточности и инфаркта миокарда у людей с ожирением. Обнаружена прямая корреляция между индексом массы тела (ИМТ) и гипертрофией левого желудочка (ГЛЖ) на моделях животных и человека. Механизм воздействия лептина на сердце, по-видимому, носит смешанный характер – через симпатическую стимуляцию, а также за счёт воздействия на кардиальные рецепторы лептина. Ещё одно важное направление в действии лептина, которое необходимо отметить, связано с повреждающим влиянием его на внутреннюю структуру сосудистой стенки. Это действие заключается в снижении релаксации артерий, усилении кальцификации сосудов и в потенцировании протромботической агрегации тромбоцитов. Все эти воздействия могут быть отнесены к факторам, способствующим развитию атеросклеротического повреждения сосудов и формированию атеросклеротической бляшки. Их значимость возрастает ещё больше у пациентов с ожирением, у которых регистрируется повышенные плазменные уровни многих протромботических факторов. Эффективному снижению этих протромботических рисков могут способствовать диетические рекомендации, направленные против ожирения.
7 Лептин и почки
Почки являются органом, где происходит основная часть процесса деградации эндогенного лептина. Наблюдалась более низкая его концентрация в почечной вене по сравнению с почечной артерией. Величина же экскреции гормона почками не зависит от концентрации лептина в крови, артериального давления, наличия или отсутствия стеноза почечной артерии. Следует отметить, что лептин в моче не обнаруживался вообще или определялся в следовых количествах, что позволило высказать предположение о внутриканальцевых превращениях гормона.
В почках (эндотелиальных клетках и мезангиуме) определяются только укороченные формы рецептора лептина obRa. Было показано, что лептин стимулирует пролиферацию эндотелиальных клеток клубочков и экспрессию TNF-β, который в свою очередь активирует механизмы, участвующие в продуцировании внеклеточного матрикса, а именно, коллагена типа IV. Увеличение отложения коллагена и повреждение эндотелиальных клеток может в конечном счёте привести к гломерулосклерозу и хронической почечной недостаточности.
Кроме того, поскольку элиминация лептина происходит, главным образом, почками, то концентрация лептина в плазме у больных хронической почечной недостаточностью заметно выше. Успешная трансплантация почек приводит к нормализации концентрации лептина.
8 Поджелудочная железа и другие отделы желудочно-кишечного тракта
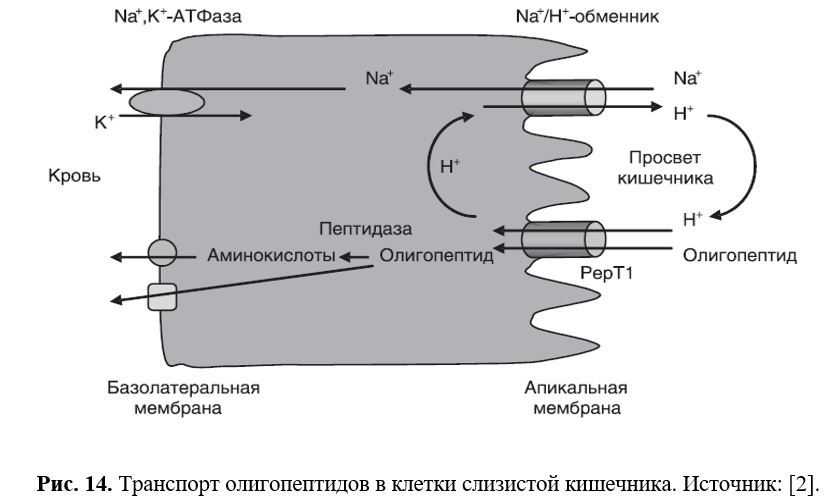
Под действием лептина отмечается подавление секреции инсулина, что осуществляется либо путём активации АТФ-зависимых калиевых каналов, либо при участии цАМФ-зависимой протеинкиназы А. Содержание инсулина в крови мышей ob/ob увеличено примерно в 100 раз, уровень гликемии повышен в 6 раз; при хроническом введении лептина концентрация инсулина и глюкозы восстанавливается. Предположительно, поступающие с периферии сигналы насыщения формируются во многом благодаря локальной экспрессии лептина в желудке. Лептин, вырабатывающийся в кишечнике, участвует в процессах всасывания нутриентов, а также в обеспечении моторики ЖКТ. Лептин, который может поступать к клеткам кишечника как через системный кровоток, так и через просвет кишечника после его секреции желудком, повышает экспрессию PepT1 на транскрипционном уровне. PepT1 – H+-пептидный котранспортёр, осуществляющий транспорт олигопептидов в кишечнике через апикальную мембрану его клеток (Рис. 15). Активность PepT1 регулируется субстратами на транскрипционном уровне, однако данные о направленности этой регуляции противоречивы.
9 Лептин и иммунная система
Тучные люди в большей степени предрасположены к воспалительным и инфекционным заболеваниям. Секретируя компоненты системы комплемента, цитокины и факторы гемопоэза, адипоциты выступают как медиаторы иммунного ответа. В условиях in vitro лептин повышает секрецию провоспалительных цитокинов из адипоцитов, активирует лимфоциты, а также ускоряет дифференцировку клеток-предшественников в гранулоциты и лимфоциты. В свою очередь, такие цитокины, как интерлейкины и фактор-ингибитор лейкемии стимулируют секрецию лептина в адипоцитах, что сопровождается снижением аппетита.
10 Лептин и СТГ
Лептин способен действовать прямо на соматотрофы гипофиза, увеличивая синтез и секрецию СТГ. Кроме того, лептин подавляет высвобождение соматостатина и стимулирует высвобождение ГРРГ гипоталамусом, что также должно способствовать секреции СТГ. Лептин служит также пермиссивным фактором для активации репродуктивной системы, которая через половые гормоны стимулирует секрецию СТГ [14]. Физиологическое значение стимулирующего действия лептина на продукцию СТГ, по-видимому, заключается в поддержании массы тела на постоянном уровне через систему обратной связи: наполнение адипоцитов жиром — повышение секреции лептина — увеличение уровня СТГ — стимуляция липолиза, ингибирование липосинтеза.
11 Лептин, тиреоидные гормоны и термогенез
Тиреоидные гормоны наряду с симпатической нервной системой, служат эффекторами в стимулирующем действии лептина на энергообмен. Интенсивность энергообмена определяется не только температурными условиями окружающей среды, но и доступностью пи-
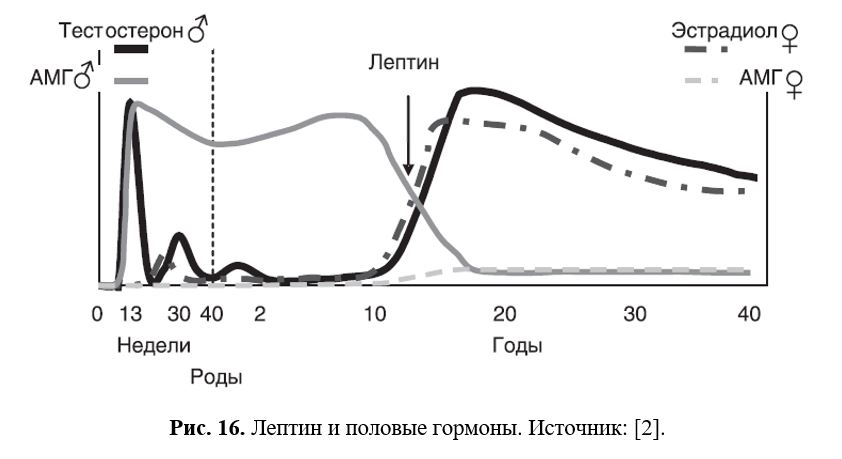
щи. Так, при голодании в тканях адаптивно снижается экспрессия дейодиназы-2, что приводит к снижению локально образующегося T3 и соответственно уменьшению интенсивности обменных процессов и сохранению питательных веществ в организме. И напротив, увеличивающиеся жировые запасы при избытке пищи стимулируют секрецию адипоцитами лептина, который через активацию симпатико-адреналовой системы и тиреоидной функции повышает термогенез, что ограничивает дальнейший рост массы тела. Однако связь между терморегуляцией и режимом питания осуществляется не только на уровне периферических тканей, но и на уровне ЦНС (Рис. 16).
12 Лептин и половая система
Пермиссивным фактором для пубертатного подъема секреции половых гормонов гонадами служит лептин (Рис. 17).
Мутация гена лептина у человека приводит к развитию ожирения и гипогонадизма. У ob/ob мышей с генетическим отсутствием лептина введение данного гормона восстанавливает фертильность. Действие лептина на половое созревание может происходить как на уровне гипоталамуса, так и на уровне половых желез. В гипоталамусе данный механизм может осуществляться через производные соединения проопиомеланокортина или через NPY. В течение менструального цикла уровень лептина постепенно нарастает на протяжении фолликулярной фазы, достигая пика в лютеиновую фазу. В норме отмечается также синхронность пульсового выброса лептина и лютеинизирующего гормона гипофиза. Во время беременности уровень лептина повышается вместе со сроком гестации в большей степени, чем это соответствует нарастанию массы тела, что предполагает развитие физиологической лептинорезистентности. После родов уровень лептина резко снижается, что может отражать энергетические затраты лактационного процесса. Хотя достаточный уровень лептина необходим для достижения фертильности, у ob/ob мышей с генетическим отсутствием лептина после наступления беременности прекращение экзогенного введения данного гормона не препятствовало дальнейшему развитию беременности. Было обнаружено, что лептин содержится в фолликулярной жидкости в тех же концентрациях, что и в сыворотке крови
Кроме того, концентрация гормона в крови после наступления половой зрелости в 2-3 раза выше у женщин, чем у мужчин. В фертильном возрасте снижается содержание лептина у мужчин и увеличивается у женщин из-за более выраженного подавляющего воздействия андрогенов на синтез лептина, чем стимулирующего – эстрогенов. В то же время, у женщин отмечается большее содержание общего жира в организме, чем у мужчин, что также сказывается на его концентрации. Подкожные адипоциты продуцируют в два раза больше лептина из расчета на клетку, чем внутриабдоминальные клетки, возможно, отчасти по причине большего объема подкожных, нежели висцеральных адипоцитов.
Существуют противоречивые данные о влиянии лептина на развитие молочной железы. С одной стороны, у мышей с недостаточностью лептина (линия ob/ob) или его рецептора (линия db/db) развитие молочной железы было минимальным, что свидетельствует о стимуляции лептином пролиферации эпителия. С другой стороны, экспрессия лептина в молочной железе подавляется лактогенными гормонами.
Интересно отметить исследования о взаимодействии лептина и кортизола. Результаты довольно неоднозначны. Суточный ритм лептина и кортизола носит противоположный характер: пик секреции лептина совпадает с минимальным уровнем кортизола. В культуре клеток коры надпочечника физиологические дозы лептина тормозят продукцию кортизола, стимулированную АКТГ [12]. А в паравентрикулярных ядрах гипоталамуса лептин, наоборот, увеличивает экспрессию кортиколиберина. По-видимому, лептин и гипоталамо-гипофизарно-надпочечниковая система находятся в реципрокных отношениях и осуществляют своё взаимодействие на различных уровнях. Значимость лептина в регуляции гипоталамо-гипофизарно-надпочечниковой активности у человека, как в норме, так и при патологии в настоящее время активно изучается.
Важно сказать и про связи с инсулином. Поскольку лептин и инсулин являются главными регуляторами энергетического гомеостаза, внимание исследователей фокусируется на их взаимоотношениях, которые до сих пор уточняются. Показано, что лептин влияет на метаболизм глюкозы, независимо от анорексигенного эффекта. Внутривенное введение лептина увеличивает продукцию глюкозы и уменьшает содержание гликогена в печени. Предполагается, что лептин влияет непосредственно на метаболизм глюкозы в печени, но не известно, противоположно ли оно действию инсулина или аналогично. Показано, что инсулин участвует в регуляции уровня лептина сыворотки. Продемонстрировано, что уровень лептина у пациентов при впервые диагностированном сахарном диабете 1 типа низок, но увеличивается после начала инсулинотерапии. Больные, получающие интенсифицированную инсулинотерапию, имеют более высокий уровень лептина, чем пациенты на стандартной инсулинотерапии. Это, вероятно, является следствием стимулирующего влияния инсулина на продукцию лептина. Другие авторы не отметили существенного влияния инсулина на сывороточную концентрацию лептина у здоровых людей. У больных сахарным диабетом 2 типа уровень лептина сыворотки выше, независимо от объема висцерального жира. Наиболее высокая лептинемия натощак наблюдается у пациентов с максимально выраженной инсулинорезистентностью. Показано, что при липодистрофии, характеризующейся отсутствием жировой ткани и крайне низким уровнем лептина, часто развивается сахарный диабет 2 типа. Описаны влияния лептина на изолированные панкреатические островки, где он блокирует базальный и стимулируемый глюкозой выброс инсулина путем воздействия на чувствительные к АТФ калиевые каналы. С другой стороны, имеются свидетельства действия лептина как медиатора выработки инсулина β-клетками. Показано, что гормон может уменьшать степень инсулинорезистентности, вероятно, не только за счет периферических, но и за счет центральных эффектов. Обсуждаются также внутриклеточные механизмы передачи сигналов, стимулируемые и лептином, и инсулином.
13 Методы тестирования уровня гормона
Уровень лептина измеряется путём стандартного анализа крови. Венозная кровь является материалом для исследования. Применяют Наиболее распространенной методикой проведения анализа является иммуноферментный анализ (ИФА), в основе которого лежит специфическая реакция антиген-антитело. Искомое вещество (лептин) обнаруживается, благодаря добавлению меченного реагента (коньюгата), который, специфически связываясь только с этим веществом (лептин), окрашивается. Интенсивность окраски пропорциональна количеству определяемого вещества в сыворотке крови. Предлагается реагент Leptin Human ELISA.
Как уже отмечалось, концентрация гормона в крови зависит от многих факторов: пола, возраста, времени суток и состояния организма, а также уровень лептина в крови коррелирует с запасами жировых депо. У грызунов концентрация гормона повышается спустя несколько часов после приема пищи, а у человека – через несколько дней (по некоторым данным, спустя 18-24 часа после ограничения поступления пищи); снижение показателей у животных и человека наблюдается в течение нескольких часов после начала голодания.
Однако концентрация циркулирующего лептина может значительно различаться среди лиц с одинаковой массой жира, что может быть следствием воздействия на экспрессию лептина инсулина, глюкокортикоидов и половых гормонов.
Как и у большинства гормонов, секреция лептина носит пульсовой характер с колебаниями в течение суток. Максимальный уровень лептина в крови отмечается после полуночи (22.00-03.00), минимальный – около полудня.
Концентрация в крови здорового человека составляет 7,5±8,5 мкг/л и 3,8±17,3 мкг/л у мужчин и женщин соответственно.
Литература
- Смирнов А.Н. Элементы эндокринной регуляции / А.Н.Смирнов — М.: ГЭОТАР — Медиа, 2008.
- Смирнов, А.Н. Эндокринная регуляция. Биохимические и физиологические аспекты: учебное пособие / А.Н. Смирнов; под ред. В.А. Ткачука — М.: ГЭОТАР-Медиа, 2009.
- Thon Mina, Hosoi Toru, Ozawa Koichiro Possible Integrative Actions of Leptin and Insulin Signaling in the Hypothalamus Targeting Energy Homeostasis, — Frontiers in Endocrinology, 2016.
- Wauman Joris, Zabeau Lennart, Tavernier Jan The Leptin Receptor Complex: Heavier Than Expected? — Frontiers in Endocrinology, 2017.
- Дедов И. И., Мельниченко Г. А., Бутрова С. А. Жировая ткань как эндокринный орган // Ожирение и метаболизм. 2006. №1.
- МакДермотт М. Секреты эндокринологии / под ред. Ю.А. Князева, М.А. Борисовой, Л.Я. Рожинской — М.: Бином, 2001.
- Романцова Т. И., Волкова Г. Е. Лептин и грелин: антагонизм и взаимодействие в регуляции энергетического обмена // Ожирение и метаболизм. 2005. №2 С.2-9.
- Чубриева С. Ю., Глухов Н. В., Зайчик A. M. Жировая ткань как эндокринный регулятор (обзор литературы) // Вестник СПбГУ. Серия 11. Медицина. 2008. №1 С.32-44.
- Кучер А.Г., Смирнов А.В., Каюков И.Г., Добронравов В.А., Яковенко А.А. Лептин – новый гормон жировой ткани: значение в развитии ожирения, патологии сердечно-сосудистой системы и почек // Нефрология. 2005. Том 9. № 1.
- Аблаев Н.Р. Лептин и его роль в организме человека // Лабораторная медицина. 1-2 (4) 2012-2013 С.30-36
- Романцова Т.И., Волкова Г.Е. Лептин и грелин: антагонизм и взаимодействие в регуляции энергетического обмена // Ожирение и метаболизм. 2005. №2.
- Коваренко М.А., Руяткина Л.А., Петрищева М.С., Бодавели О.В. Лептин: физиологические и патологические аспекты действия // Вестник НГУ. Серия: биология, клиническая медицина. — 2003. — Т.: 1, Вып.: 1. — С. 59-74.
- Чубриева С.Ю., Глухов Н.В., Зайчик А.М. Жировая ткань как эндокринный регулятор (обзор литературы) // Вестник Санкт-Петербургского университета. 2008. Сер.11. Вып.1.
- Верин, В. К. Гормоны и их эффекты: справочник / В. К. Верин, В. В. Иванов. – СПб.: Фолиант, 2012.
- Физиология эндокринной системы. http://biomedia.pro/physiology/endocrinology/
- J. Larry Jameson Harrison’s Endocrinology / Derived from Harrison’s Principles of Internal Medicine, 17th Edition, 2010.
Reference
- Smirnov A.N. Elements of endocrine regulation / A.N. Smirnov — M .: GEOTAR — Me-dia, 2008.
- Smirnov, A.N. Endocrine regulation. Biochemical and physiological aspects: study guide / A.N. Smirnov; by ed. V.A. Tkachuka — Moscow: GEOTAR-Media, 2009.
- Thon Mina, Hosoi Toru, Torting Energy Homeostasis, — Frontiers in Endocrinology, 2016.
- Wauman Joris, Zabeau Lennart, Tavernier Jan The Leptin Receptor Complex: Heavier Than Expected? — Frontiers in Endocrinology, 2017.
- Dedov I. I., Melnichenko G. A., Butrova S. A. Adipose tissue as an endocrine organ // Obesity and metabolism. 2006. №1.
- McDermott M. Secrets of Endocrinology / ed. Yu.A. Knyazev, M.A. Borisova, L.Ya. Rozhinsky — M .: Binom, 2001.
- Romantsova T.I., Volkova G.E. Leptin and ghrelin: antagonism and interaction in the regulation of energy metabolism // Obesity and Metabolism. 2005. №2 С.2-9.
- Chubrieva S. Yu., Glukhov N. V., Zaichik A. M. Adipose tissue as an endocrine regulator (literature review) // SPbSU Bulletin. Series 11. Medicine. 2008. №1 С.32-44.
- Kucher A.G., Smirnov A.V., Kayukov I.G., Dobronravov V.A., Yakovenko A.A. Leptin — a new hormone of adipose tissue: importance in the development of obesity, pathology of the cardiovascular system and kidneys // Nephrology. 2005. Volume 9. № 1.
- Ablaev N.R. Leptin and its role in the human body // Laboratory medicine. 1-2 (4) 2012-2013 P.30-36
- Romantsova T.I., Volkova G.E. Leptin and ghrelin: antagonism and interaction in the regulation of energy metabolism // Obesity and metabolism. 2005. №2.
- Kovarenko M.A., Ruyatkina L.A., Petrishcheva M.S., Bodaveli O.V. Leptin: physiological and pathological aspects of action // Bulletin of the NSU. Series: biology, clinical medicine. — 2003. — T .: 1, Issue: 1. — p. 59-74.
- Chubrieva S.Yu., Glukhov N.V., Zaychik A.M. Adipose tissue as an endocrine regulator (literature review) // Bulletin of St. Petersburg University. 2008. Ser.11. Issue 1.
- Verin, V. K. Hormones and their effects: a handbook / V. K. Verin, V. V. Ivanov. — SPb .: Foliant, 2012.
- The physiology of the endocrine system. http://biomedia.pro/physiology/endocrinology/
- J. Larry Jameson Harrison’s Endocrinology / Derived from Harrison’s Principles of Internal Medicine, 17th Edition, 2010.